Ничего лишнего
Я думаю, что человек, пишущий эти статьи, делает это в целях просвещения как можно большего количества людей и он не поставит мне в вину то, что я размещаю их в своем дневнике. Ибо у меня нет других целей.
читать дальшеF134
--------------------------------------------------------------------------------
В генетике многоклеточных организмов нельзя пройти мимо загадки целостности организма. Вероятно, нужно искать не способ кодирования каких-то дискретностей, а способ записи информации о неразрывной трехмерной (а с учетом времени — четырехмерной) анатомии всего организма.
«Из открытий Менделя, апробированных победным развитием генетики ... вытекает бесспорный принцип дискретности наследственной детерминации признаков. Однако бесспорно и то, что эмбриология не имеет оснований говорить о независимом возникновении и развитии признаков в онтогенезе. Нормальный онтогенез — это совокупность процессов изменения состояния целостности. Что означает противоречивость двух бесспорных утверждений?» [Токин, 1987]
Загадка целостности организма в индивидуальном развитии многократно приводила исследователей к идее биополя, управляющего пространственной организацией живой материи. Иногда авторы не говорили о биополе напрямую, но логикой фактов подводили к мысли о его существовании.
Термин «биополе» используется не только в биологии развития, где его применяют к внутриорганизменным процессам полевой природы. Термин «биополе» используется также биофизиками, экстрасенсами и др., но уже в ином смысле — применительно к процессам полевой природы в пространстве, окружающем организм. Мы будем рассматривать только процессы, протекающие внутри организмов.
Как отмечалось ранее, иногда формы одного животного (контур рыбы, кость птицы и т.п.) удается точно совместить с формами животного другого биологического вида, если подвергнуть изображение простым деформациям — плавному растяжению, сжатию или др. [Thomрson, 1942]. Здесь тоже видны явления полевой природы, проглядывают законы, которые нельзя выразить в одномерном или даже в двумерном пространстве. Глазам исследователей предстает явная передача от предков к далеким потомкам не только биохимических процессов, но и пространственного образа, способного плавно деформироваться в ходе эволюции. Формирование пространственного образа подразумевает протекание неких пространственно-организованных событий, наводя на мысль о биополе.
Обстоятельный (хотя далеко не полный) обзор разработок проблемы биополя приведен в работе Токина от 1987 года.
Довольно развитые представления о биополе изложил П. Вейс, уподобивший его системе векторов. Представления Вейса явились результатом его исследований по регенерации органов. По Вейсу, поле материально, но его природа не может быть сведена к химической, так как, по его мнению, химические факторы способны лишь активировать, но не формировать поле.
Н. К. Кольцов на основе изучения процессов в ооцитах и яйцах создал представление об управляющем воздействии на зародыш ряда «силовых» полей в виде разностей электрических, химических, температурных, гравитационных, диффузионных и других потенциалов. В ходе развития силовое поле зародыша усложняется, дифференцируется, но остается единым.
Близкую к взглядам Н. К. Кольцова трактовку морфогенетических процессов предложил в 1958 г. Б. Вейсберг, который на основе изучения колебаний электрических потенциалов у миксомицетов создал представление о «колебательных полях».
Идейную близость к подобным взглядам можно заметить в теории «аксиальных градиен-тов» американского биолога Ч. Чайлда.
Интересные этапы проходила концепция биополя, разрабатывавшаяся Александром Гавриловичем Гурвичем [Гурвич, 1944; Любищев, Гурвич, 1998]. Отметим некоторые из его тезисов, хотя подвергнутые критике, но, как мы увидим далее, хорошо отражающие реальность:
•поведение элементов всех микроморфогенетических процессов управляется единым фактором — «преформированной морфой», задающей еще не существующую структуру и определяющей окончательную конфигурацию зачатка;
•каждая клетка имеет собственное поле, действие которого выходит за пределы клетки; при делении клетки делится и ее поле; клетки оказывают своими полями влияние друг на друга; по мере удаления влияние поля быстро ослабевает;
•в ходе индивидуального развития происходит эволюция «поля целого»; то целое, что существует в данный момент, определяет ход дальнейшего развития.
Даже краткий обзор полевых концепций в биологии и связанных с ними исследований занял бы много места. Идею существования в организме некого биополя, участвующего в управлении морфогенезом, в разное время и с разных позиций развивали, например, И. А. Аршавский, Л. В. Белоусов, Г. Р. де Бер, Т. Бовери, Б. Брайент, Л. Вольперт, Дж. С. Гексли, Е. Гиено, В. В. Исаева, Г. П. Короткова, А. А. Любищев, В. М. Маресин, Е. В. Преснов, П. Г. Светлов, К. Уоддингтон, П. Френч, Д. С. Чернавский, Г. Шпеман, Т. Ямада и другие ученые. Их взгляды базировались на глубоких исследованиях, оставивших заметный след в науке.
К сожалению, сильнее оказалось отрицательное влияние многочисленных энтузиастов, создавших вокруг проблемы ажиотаж, но не приблизивших понимание природы явления. Идея биополя оказалась серьезно дискредитированной ими, так что сегодня биологи упоминают ее преимущественно в уничижительном тоне.
Сегодня ясно, что исследования нескольких десятилетий принципиально не могли раскрыть природу внутреннего биополя организма, поскольку этому должно было предшествовать фундаментальное открытие в иной области. Но изложим события по порядку.
Идея биополя близка к идее волнового информационного взаимодействия организма с генами. Мысль о том, что формирование многоклеточного организма связано с передачей информации волновыми полями, имеет немалую предысторию, она высказывалась рядом исследователей. Например, А. Н. Мосолов [1980] предположил, что на гены воздействует акустическое поле. Но акустические (и электромагнитные) колебания, которые могли бы взаимодействовать с генами (т.е. имели бы протяженность фронта волны, сопоставимую с размером гена) — такие колебания очень сильно затухают в биологических тканях, отчего бессмысленно говорить об их распространении более чем на миллиметры. В то же время, чтобы связать формирование организма с волновым управлением генами, нужны волны, способные без чрезмерного затухания распространяться по всему организму.
Ситуация казалась безвыходной, пока не были открыты волны совершенно новой природы. Стремясь создать удобную для экспериментов упрощенную модель цикла Кребса — цепочки реакций, питающих наш организм энергией, Борис Павлович Белоусов открыл в 1951 году колебательную химическую реакцию. Наблюдать такую реакцию очень легко; достаточно смешать в пробирке 10мл водного раствора веществ:
•лимонная кислота — 2,00 г
•сульфат церия — 0,16 г
•бромат калия — 0,20 г
•серная кислота (1:3) — 2,0 мл
•воды до общего объема — 10,0 мл.
При комнатной температуре в пробирке возникают периодические изменения цвета «желтый-бесцветный» с периодом порядка десятков секунд. Более контрастное изменение цвета «синий-красный» происходит при добавлении к смеси железофенантролина в качестве индикатора. Нагревание ускоряет смену цветов. Постепенное расходование ингредиентов ведет к затуханию процесса, а прибавление израсходованного вещества возобновляет его.
Однако химические журналы отказались публиковать сообщение о колебательной реакции, считая статью блефом. Лишь в 1959 г. появилось короткое сообщение в журнале по радиационной медицине, который химики не читают. В 1970 г. Б. П. Белоусов умер, так и не сумев донести открытие до научной общественности . А в 1980 году за это открытие была присуждена Государственная премия. С легкой руки Ильи Романовича Пригожина — президента Бельгийской Королевской академии наук — открытие Б. П. Белоусова было названо самым выдающимся экспериментом ХХ века.
Реакция БЖ К этому времени А. М. Жаботинский и А. Н. Заикин установили, что колебательные реакции способны распространяться в соответствующей среде в виде волн. На фото запечатлен один из моментов распространения волн химических реакций в тонком слое жидкости, налитой на дно плоского сосуда.
К этому времени А. М. Жаботинский и А. Н. Заикин установили, что колебательные реакции способны распространяться в соответствующей среде в виде волн. На фото запечатлен один из моментов распространения волн химических реакций в тонком слое жидкости, налитой на дно плоского сосуда.
Такие волны назвали волнами Белоусова-Жаботинского (БЖ). Они, как лесной пожар, черпают энергию из среды, по которой распространяются, отчего, даже при очень серьезных потерях энергии, способны распространяться на неограниченно большие расстояния.
Открытие колебательных реакций, как часто бывает, стало результатом удачного стечения обстоятельств. Если бы период изменения цвета в пробирке измерялся сотыми долями секунды или, наоборот, многими часами и днями, то явление осталось бы незамеченным. Между тем, есть основания считать, что подобные реакции с сильно отличающимися временными характеристиками реально существуют и достаточно распространены. В этом смысле привлекает внимание причудливый узор разрезанного агата. Похоже, что он изображает застывшие волны, связанные с химическими процессами, но протекавшие во много раз медленнее, чем в случае волн БЖ.
Среди многообразия химических реакций есть и такие, что распространяются с очень высокими скоростями, например, взрывы, детонация в двигателях и др. Для скорости продвижения химических волн важен механизм их распространения. Если в его основе лежит диффузия молекул, то скорость волны будет зависеть от размера молекул, температуры и вязкости среды. Однако агентами, инициирующими реакцию в соседних зонах, могут быть также электроны, протоны, кванты излучения, рождающиеся в ходе реакции, и они определят другие, гораздо более высокие скорости ее распространения.
Это дает основания предполагать, что среди большого числа реакций, протекающих в протоплазме клеток, существуют и такие, что распространяются в виде волн, хотя они, из-за временных характеристик или по другим причинам, пока напрямую не обнаружены. Неслучайно Б. П. Белоусов открыл колебательные реакции, пытаясь моделировать едва ли не самую распространенную в живой природе цепочку биохимических процессов — цикл Кребса.
Один из исследователей [Pohl, 1983] на основании двух независимых экспериментальных методик выяснил, что, по-видимому, волны химических реакций возникают в протоплазме каждой нормальной эукариотической клетки. Тем самым, подкрепилось предположение, что волны, переносящие информацию при формировании многоклеточного организма, являются волнами химических реакций.
Веские основания искать в организме химические колебательные реакции дает картина переменной электрической активности тканей, которую нельзя свести только к активности нейронов и мышц. Например, описано появление особой компоненты колебаний электрического потенциала во время восстановления (регенерации) сломанной кости лапы кролика [Бутуханов, Ипполитова, 1982].
F135
--------------------------------------------------------------------------------
Волны химических реакций, среди которых особую известность получили волны БЖ, оказались неожиданными не только по своей природе. Как явствовало уже из первых осциллограмм, полученных Б. П. Белоусовым, они относились к особому классу — к классу уединенных волн или солитонов.
В настоящее время студенты ВУЗов изучают волновые процессы на примере синусоидальных колебаний, и лишь немногие из них получают теоретические представления о солитонах, которые являются единственным типом волн физиологических процессов. Уже изучая практические предметы, биологи и медики, не зная теории, сталкиваются с солитонами в виде нервных импульсов, волн сокращений миокарда, разрядов электрического ската, волн перистальтики, волн нервного возбуждения в тканях мозга и т.д. Поэтому есть смысл сказать несколько слов о солитонах вообще.
Физики и математики делят колебания на линейные и нелинейные. Теория линейных колебаний создана в XVIII-XIX веках и лежит в основе аппарата современной математической физики. Синусоидальные колебания — это линейные колебания. Систематическая теория нелинейных колебаний и волн — теория солитонов — появилась во второй половине XX века. Познакомиться с ней можно по работам [Филиппов, 1986; Додд и др., 1988; Ньюэлл, 1989] и многим другим.
Синусоидальные колебания в чистом виде — наиболее простые колебания, колебания одной определенной частоты, строго симметричные относительно исходного уровня. Для прохождения синусоидального колебания по каналам связи требуется минимальная ширина полосы частот. Все это обусловило широкое использование синусоидальных колебаний в технике, их почетное место в учебных программах.
Солитоны, как правило, распространяются в нелинейных средах, что существенно влияет на их особенности.
1.Во-первых, нелинейность среды определяет некоторую оптимальную амплитуду солитона, от которой ему, в отличие от синусоидальных колебаний, труднее отклоняться и в меньшую, и в большую стороны.
2.Во-вторых, нелинейность среды создает для него некую возможность обмена одних параметров на другие, скажем, амплитуды на форму. Последнее придает солитону способность распространяться без заметных изменений амплитуды на гораздо большие расстояния, чем мы ожидали бы по опыту работы с синусоидальными волнами.
3.Наконец, в-третьих, нельзя не отметить, что нелинейность среды распространения солитонов часто определяется активностью среды и это особенно характерно для биологических тканей. В таких случаях размах и форма проходящего солитона имеют повышенную стабильность, потому что по мере движения солитона среда подпитывает его своей энергией.
Пожалуй, наиболее существенным свойством солитонов, резко отличающим их от синусоидальных колебаний, является независимость друг от друга. У синусоиды нельзя отделить одну волну от других или сдвинуть относительно других, изменить размах. Это сразу превратит синусоиду определенной частоты в нечто иное — в сложную совокупность разночастотных синусоидальных колебаний. Ничего похожего не грозит солитонам. Каждый из них можно, например, смещать во времени, и это не изменит ни его основных параметров, ни параметров других солитонов.
Соответственно, по отношению к солитонам нужно с большой осторожностью говорить о частоте. Они могут следовать на одинаковом расстоянии друг от друга (тогда обретает смысл понятие частоты следования), но могут от волны к волне резко изменять интервал или вовсе следовать через хаотически меняющиеся промежутки времени. В биологических процессах встречаются все эти случаи.
Солитон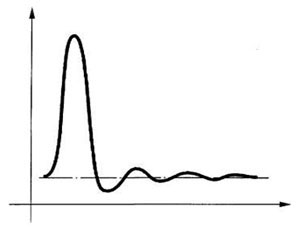 Типичная форма солитона показана на рисунке справа. Она может существенно изменяться в зависимости от природы процесса и параметров среды. Например, затухающие колебания за задним фронтом волны возникают только при существенном ухудшении условий прохождения высоких частот.
Типичная форма солитона показана на рисунке справа. Она может существенно изменяться в зависимости от природы процесса и параметров среды. Например, затухающие колебания за задним фронтом волны возникают только при существенном ухудшении условий прохождения высоких частот.
Часто солитоны обладают большой скважностью, т.е. расстояние (во времени) между соседними волнами оказывается во много раз большим, чем длительность самого колебания. В таких случаях энергия колебаний сосредотачивается на гораздо более высоких частотах, чем частота следования волн.
При резком различии скоростей процессов, определяющих передний и задний фронты волны, задний фронт может превратиться в медленно спадающую кривую, что придает солитону вид одиночного скачка параметра с последующим длительным (по сравнению с продолжительностью переднего фронта) возвращением в исходное состояние. Такую разновидность солитонов иногда называют кинками. Волны Белоусова-Жаботинского имеют именно такую форму — у них процессы, определяющие задний фронт волны, протекают намного медленнее, чем процессы, от которых зависит крутизна переднего фронта.
Способность солитонов следовать друг за другом через произвольные интервалы времени открывает для них большие возможности в части переноса информации. Кроме того, солитон, по сравнению с синусоидальными колебаниями, очень устойчив против возмущений, вносимых неоднородностью среды распространения и внешними помехами [Захаров, Шабат, 1962]. Эти свойства приобретают особую значимость при переносе информации в такой неоднородной и слабо защищенной от внешних воздействий среде, как живой организм.
Многие математики не согласны называть волны химических реакций солитонами. По их мнению, химические волны и солитоны — это разные понятия, разные категории объектов. Но в таком возражении проявляется лишь желание математиков подогнать понятие солитона под класс объектов, которые они научились описывать. Волны химических реакций они пока описывать не умеют. Между тем, многие специалисты вполне логично рассматривают солитон как нелинейное устойчивое локализованное возбуждение (например, [Шахбазов и др., 1996]), а такая трактовка охватывает и волны химических реакций.
Оптика нового типа
«Возможно такое, что вам и не снилось... Я буду рассказывать, как устроена Природа, и, если вам не понравится, как она устроена, это будет мешать вашему пониманию. Физики научились решать эту проблему: они поняли, что, нравится им теория или нет — неважно. Важно другое — дает ли теория предсказания, которые согласуются с экспериментом. Тут не имеет значения, хороша ли теория с философской точки зрения, легка ли для понимания, безупречна ли с точки зрения здравого смысла. Квантовая электродинамика дает совершенно абсурдное с точки зрения здравого смысла описание Природы. Но оно полностью соответствует эксперименту. Так что я надеюсь, что вы сможете принять Природу такой, какая она есть — абсурдной».
Фейнман, 1988
Сам факт осциллографической регистрации Б. П. Белоусовым химических колебаний показал, что в ходе реакции скачкообразно изменяется окислительно-восстановительный потенциал среды. Как оказалось, химической волне сопутствует скачок электрического потенциала порядка десятых или сотых долей вольта. Но есть ли основания считать, что подобные волны возникают в живых организмах?
Непосредственная регистрация в организме колебаний с ожидаемыми параметрами встречает серьезные технические трудности. Косвенные данные говорят, что длительность переднего фронта ожидаемых волн имеет временную протяженность порядка десятков пикосекунд. Это значит, что основная энергия колебаний сосредоточена в области десятков гигагерц. К тому же, мощность колебаний в расчете на одну клетку очень мала. Самые современные осциллографы еще не способны зарегистрировать подобные волны, показать их форму.
Учитывая, что увидеть осциллограмму структурогенных волн пока не удается, Герберт Пол заменил прямую регистрацию косвенными исследованиями на основе микродиэлектрофореза. В раствор, окружающий выделенную живую клетку, Пол ввел мельчайшие гранулы диэлектрика и наблюдал под микроскопом их поведение. В зависимости от того, у какого вещества — у жидкости или у гранул — была выше диэлектрическая проницаемость, частицы двигались к клетке, либо в противоположную сторону [Pohl, 1983]. Это четко указывало на существование в клетке переменного электрического поля. Но оставалось неясным, привязаны ли электрические колебания к поверхности клеток или же они протекают в объеме цитоплазмы?
На этот вопрос ответила другая серия опытов. Пол создал вокруг взвешенной в жидкости клетки вращающееся электрическое поле, и увидел, что клетка тоже стала вращаться, хотя и с меньшей скоростью. Более того, при изменении скорости вращения поля клетка обнаружила резонансные свойства — на определенных частотах она резко ускоряла свое вращение. Этот эффект, названный клеточным спиновым резонансом, противоречил предположению об электрических колебаниях на так называемой мембране, но говорил в пользу химических волновых процессов в протоплазме клетки.
Что произойдет при движении волны химических реакций в протоплазме клетки, когда фронт волны пересечет клеточное ядро?
В кольцевой зоне оболочки клеточного ядра, где в какой-то момент расположен скачок окислительно-восстановительного потенциала, возникнет очень высокая напряженность электрического поля. Скачок потенциала порядка десятых долей вольта воздействует на поверхностный слой (ошибочно называемый мембраной) общей толщиной всего 14 нм и создает в нем напряженность поля до 200 кВ/см. Вследствие этого в зоне соприкосновения фронта волны с ядерной оболочкой должно произойти интенсивное электрострикционное сжатие диэлектрического материала оболочки. Согласно расчетам [Артюшин, Барбараш, 1985], если бы отсутствовало гидравлическое сопротивление внутриядерной жидкости, то деформация оболочки достигла бы 10% диаметра ядра.
Быстрое перемещение кольцевой зоны деформаций по оболочке ядра порождает во внутриядерной жидкости акустические колебания. Скорости волн различной природы обычно не совпадают. Поэтому на границе раздела — на оболочке ядра, где энергия химических волн переходит в энергию акустических, происходит преломление волнового поля. Оболочка ядра является, в первом приближении, сферой. Явления, связанные с преломлением волнового поля на сферической поверхности, досконально изучены оптиками — такая поверхность приобретает свойства линзы. Конечно, того, что принято называть линзой, здесь нет. Но это не меняет результата. Возможно, следует ввести новый термин — линза-мембрана, мембролинза или что-либо в этом роде.
В обычной оптике преломляющиеся волны не изменяют свою природу в момент преломления, т.е. при переходе из области с одной скоростью распространения в область с другой скоростью. В обычной оптике изменение скорости распространения объясняется не изменением природы волнового поля, а изменением свойств среды распространения. Такую оптику можно условно назвать гомоволновой (homos — одинаковый). В нашем же случае изменение скорости вызывается именно изменением природы волнового поля. Здесь принципиально важно, что волны одной природы при достижении некой преобразующей пленки превращаются (передают свою энергию) в волны иной природы. Оптику этого неизвестного ранее типа можно назвать гетероволновой оптикой (heteros — иной, другой).
Гетероволновая оптика обладает рядом особенностей. Поскольку в ней могут сочетаться волновые процессы с сильно различающимися скоростями распространения, то оказываются достижимыми очень высокие коэффициенты преломления, а это позволяет при прочих равных условиях резко сократить количество компонентов оптической схемы и уменьшить аберрации. Если пленка-преобразователь работает на основе необратимого физического явления (например, в оболочке клеточного ядра возникает необратимая электрострикция, а не обратный пьезоэлектрический эффект), то из высокого коэффициента преломления не следуют, как это обычно бывает, высокие потери на отражение (так как не индуцируется противоволна). В гетероволновой оптике фокусирующими элементами служат не линзы, а тонкие пленки, из-за чего такая оптика имеет очень низкую материалоемкость.
Таким образом, оболочка ядра, действуя как линза гетероволновой оптики, преобразует химические волны протоплазмы в акустические колебания внутриядерной жидкости.
Термин «оптика» имеет два толкования — узкое и расширенное. В узком понимании это наука о том, что происходит с лучами видимой части спектра электромагнитных колебаний при их отражении, преломлении, поглощении и т.д. В расширенном понимании оптика охватывает процессы, в которых могут участвовать не только электромагнитные волны видимой части спектра, но и другие волны и объекты, поведение которых описывается сходными законами.
В расширенном понимании для разных конкретных задач термин «оптика» обычно дополняется уточняющим словом — «инфракрасная оптика», «рентгеновская оптика», «акустическая оптика», «электронная оптика», «оптика протонов» и т.д. Взаимодействие химических волн протоплазмы с клеточным ядром тоже описывается законами оптики, что дает основания отнести его к оптике в расширенном понимании. Соответственно, и здесь термин требует уточнения. Поскольку процессы связаны с ядром клетки, данный раздел оптики уместно назвать кариооптикой или сокращенно — К-оптикой.
Кариооптика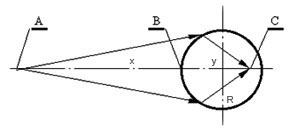 Представим себе, как показано на рисунке, что точка A относится к химическому волновому полю организма, т.е. находится вне ядра, а точка C является проекцией точки A в акустическом поле ядра. Эти точки расположатся на прямой, проходящей через центр ядра, и будут разделены ядерной оболочкой.
Представим себе, как показано на рисунке, что точка A относится к химическому волновому полю организма, т.е. находится вне ядра, а точка C является проекцией точки A в акустическом поле ядра. Эти точки расположатся на прямой, проходящей через центр ядра, и будут разделены ядерной оболочкой.
Обозначим: R — радиус ядра; x — расстояние между точкой A и ближайшей точкой В на поверхности ядра; y — расстояние от точки В до точки фокусировки С; n — отношение скоростей распространения химических и акустических волн (коэффициент преломления).
Тогда, на основании формулы нулевого инварианта Аббе [Ландсберг, 1976] получим:
nRx
y= —————— ( т.е. у = nRx/(n-1)x + R )
(n-1)x + R
Анализ этой формулы, с учетом реальных размеров клеток животных и их ядер, показывает, что при n ≥ 2.7, все пространство организма, как бы велик он ни был, проецируется внутрь ядра каждой клетки [Артюшин, Барбараш, 1985]. Такой характер проекции не нарушается при изменениях размеров ядер.
Здесь мы столкнулись с явлениями, относящимися к химии, физике диэлектриков, теории нелинейных колебаний, оптике, биологии, а вовсе не к медицине. Именно поэтому до тех пор, пока проблемами здоровья человека будут заниматься именно медики, эту область естествознания будет окутывать полный мрак.
F136
--------------------------------------------------------------------------------
«Продвижение науки вперед часто происходит тогда, когда перед нами раскрывается неведомая доселе сторона вещей, что обусловлено не столько применением какого-то нового метода, сколько рассмотрением объектов под другим углом зрения».
©Jacob, 1977
На основе открытия колебательных реакций и волн Белоусова-Жаботинского, еще в 1982 году была разработана концепция структурогенеза (в дальнейшем — КСГ), включившая в себя впервые сформулированную идею гетероволновой оптики и объяснившая на такой основе принципы генетического кодирования структуры многоклеточного организма.
Очень важной для этой работы была исходная мысль Б. П. Белоусова о сходстве волновых химических реакций с процессами в живом организме. Такой подход представил известные биологические факты в новом свете — явления и закономерности, ранее казавшиеся несвязанными между собой, выстроились в четкую систему.
Согласно КСГ, в протоплазме клеток многоклеточного организма периодически самовозбуждаются волны химических реакций, пробегающие по всему организму и потому несущие информацию о его сиюминутном строении.
В протоплазме клеток бодрствующего животного такие волны маломощны и относительно разрозненны — по степени упорядоченности их можно сравнить с мелкой рябью на поверхности воды. Роль таких волн ограничивается передачей информации на малые расстояния — порядка десятков клеток.
Однако, кроме того, в определенной фазе сна, при заторможенности скелетных мышц, в паузах ритмики сердца и легких, кратковременно возникают мощные пакеты химических волн такой же природы, охватывающие весь организм. При всем их сходстве с волнами БЖ, эти волны нельзя отождествлять — у них разный химизм, разные механизмы распространения и сильно отличающиеся количественные характеристики.
Волны БЖ распространяются в растворе благодаря диффузии молекул, что определяет невысокие скорости продвижения волны — порядка миллиметров в секунду. Но диффузия активных молекул — не единственный фактор, способный инициировать реакцию в растворе. Хотя химизм структурогенных волн пока непонятен (вспомним, что от открытия Г. Менделем генов до выяснения их химической природы прошло около 90 лет!), есть веские основания предполагать, что продвижение фронта структурогенной волны обеспечивают не диффундирующие молекулы, а более быстрые частицы, обладающие, к тому же, очень коротким пробегом.
Это заставляет вспомнить об открытом А. Г. Гурвичем слабом ультрафиолетовом излучении живых клеток, которое он назвал митогенетическим. Название точно отразило подмеченную ученым связь интенсивности излучения с делением клеток (митозами), т.е. с процессами развития. Излучение универсально по отношению к живым клеткам разных типов — оно усиливается при стрессовом состоянии клетки и постепенно затухает после ее гибели. Все это заставляет думать, что УФ-кванты порождаются биохимическими реакциями, в норме свойственными клетке.
Обладая скоростью света, УФ-кванты должны проходить расстояние между последовательно вступающими в реакцию молекулами намного быстрее чем диффундирующие атомы, молекулы или радикалы. Если химическая реакция, генерирующая митогенетическое излучение, проходит по раствору в виде волны, то от такой волны нужно ожидать гораздо более высокой скорости распространения по сравнению с уже знакомыми нам волнами БЖ. Вероятно, основные потери времени приходятся не на перемещение УФ-квантов в пространстве, а на их реакцию с субстратом, т.е. на паузу между поглощением молекулой кванта и генерированием новой порции квантов.
Выше было отмечено, что проекция многоклеточного организма на хроматин ядра имеет место при коэффициенте преломления порядка 2,7 и более. Скорость акустических волн внутри клеточного ядра близка к скорости звука в воде, т.е. около 1,5 км/с. Следовательно, нужно ожидать скорости распространения химических волн порядка 1,5 х 2,7 = 4,05 км/с и выше.
Такая скорость распространения структурогенных химических волн на несколько порядков выше скорости известных волн БЖ, что как раз и говорит в пользу распространения волн в протоплазме не за счет диффузии «горячих» молекул, а благодаря упомянутым квантам ультрафиолетового излучения.
У прокариот, использующих чисто химические способы управления активностью генов, включение и выключение транскрипции осуществляют специфичные регуляторные белки. Особенности геномов эукариот заставляют думать, что их способы управления транскрипцией принципиально отличаются от такого варианта.
«В типичной эукариотической клетке ... транскрибируется только 7% всех последовательностей ДНК. Весьма маловероятно, чтобы остальные 93% ДНК были заблокированы десятками тысяч различных высокоспециализированных белков-репрессоров. Из соображений здравого смысла следует, что клетки высших организмов должны использовать вместо этого какие-то общие механизмы репрессии генов».
©Албертс и др., 1986
Действительно, эукариоты выработали общий механизм выключения активности генов, использующий намотку ДНК на нуклеосомы, что препятствует контакту ДНК-зависимых РНК-полимераз с достаточно длинным участком ДНК. Соответственно, эукариоты применили принципиально новый способ активирования генов, резко отличающийся от прокариотического. Основой нового механизма управления геномом стало использование волновых полей. Однако отказ от чисто химических способов управления генами не был стопроцентным — во многих случаях они используются эукариотами как дополнение к новому, физическому способу управления.
Практика акупунктуры распространила представление о существовании в организме неких «активных точек» и зон. Мы не будем эксплуатировать эту выгодную ситуацию. И без того ясно, что разные зоны организма — мышцы, костная ткань, спинномозговая жидкость и др. — настолько отличаются по своим свойствам, что химическое автоволновое поле этих зон принципиально не может быть всюду одинаковым, в том числе — по плотности энергии колебаний. Следовательно, нужно исходить из существования в организме зон с повышенной и пониженной концентрацией энергии химических волн.
Для реализации волнового механизма управления активностью геновγ , природе пришлось снабдить геном механизмом, реагирующим на акустические колебания. Это достигнуто с помощью остроумного решения. Из белков-гистонов, обладающих повышенной основностью (щелочностью — прим. ред.)α, были созданы крупные гранулы — нуклеосомыβ, которые, благодаря основности, хорошо «прилипают» к нити дезоксирибонуклеиновой кислоты (ДНК). В спокойной среде нуклеосомы с намотанной на них нитью ДНК сами собой собираются в крупную спираль, названную соленоидом. Находясь в соленоиде, ДНК непригодна для перезаписи с нее информации на РНК, т.е. для транскрипции. Только когда акустические колебания нуклеоплазмы разрушат структуру соленоида, оторвут нить ДНК от «прилипших» нуклеосом, — вот тогда к ДНК может подойти фермент РНК-полимераза, выполняющий синтез РНК. Затем по информации РНК особые молекулярные структуры — рибосомы — соединяют нужные аминокислоты в цепочки будущих белков.
, природе пришлось снабдить геном механизмом, реагирующим на акустические колебания. Это достигнуто с помощью остроумного решения. Из белков-гистонов, обладающих повышенной основностью (щелочностью — прим. ред.)α, были созданы крупные гранулы — нуклеосомыβ, которые, благодаря основности, хорошо «прилипают» к нити дезоксирибонуклеиновой кислоты (ДНК). В спокойной среде нуклеосомы с намотанной на них нитью ДНК сами собой собираются в крупную спираль, названную соленоидом. Находясь в соленоиде, ДНК непригодна для перезаписи с нее информации на РНК, т.е. для транскрипции. Только когда акустические колебания нуклеоплазмы разрушат структуру соленоида, оторвут нить ДНК от «прилипших» нуклеосом, — вот тогда к ДНК может подойти фермент РНК-полимераза, выполняющий синтез РНК. Затем по информации РНК особые молекулярные структуры — рибосомы — соединяют нужные аминокислоты в цепочки будущих белков.
[Определенный интерес в этой связи представляет собой высказывание Э. Ревича, сделанное им еще в 1986 году, цитируем:
«Первичной характеристикой рака (т.е. его первопричиной) является спонтанное образование патологических гистоновα. Гистоны являются важнейшими элементами во всей биологии. Из гистоновα образуются гистосомы, представляющие собой сферические образования из щелочных аминокислот связанные с другими аминокислотами. Все это вместе взятое составляет основу ядерного материала каждой клетки. Связывающиеся друг с другом гистосомы образуют нуклеосомыβ, а связывающиеся друг с другом нуклеосомы образуют геныγ. Таким образом, главным причинным фактором всех раков являются патологические гистоныα, из которых формируются патологические гистосомы. Всякий процесс в организме — нормальный или патологический — запускается конкретным геномγ.
Всякое заболевание подразумевает присутствие инородных гистосом. Если инородные гистосомы возникают эндогенным путем (т.е. внутри самого организма), то это называется изопаразитом. Иными словами, это примерно то же самое, как быть атакованным представителем другого биологического вида, но только этот интервент появился не извне, а образовался в собственном теле в результате мутации гистоновα, затем гистосом, затем нуклеосомβ и затем геновγ.
Итак, второй важнейший момент: рак — это изопаразит, возникший в результате повреждения или дефекта в гистонахα, которого можно образно представлять себе в виде представителя примитивного биологического вида, размножающегося путем формирования и распространения кист. Иными словами, рак — это спонтанное возникновение правовращающих аминокислот».
Довольно примечательно то, что Ревич и понятия не имел ни о какой КСГ, хотя о волновом механизме управления активностью генов, скорее всего, догадывался.]
Картина химического автоволнового поля организма закономерно отображается гетероволновой оптикой ядра каждой клетки — К-оптикой — в акустическом поле ядра. Каждой зоне повышенной плотности энергии волнового поля организма отвечает микроскопическая зона повышенной интенсивности акустических волн в ядре. Когда плотность акустической энергии превышает определенный порог, акустические колебания отрывают спираль ДНК от нуклеосом, т.е. разрыхляют хроматин, дают возможность ферментам подойти к расположенному в данном месте гену и тем создают условия для его транскрибирования. Так активные зоны химического волнового поля организма определяют в каждой клетке активность тех или иных генов.
[Хотя это не имеет прямого отношения к освещаемой теме, вероятно, все же имеет смысл вспомнить хорошо известный многим прибор Витафон, терапевтический эффект которого основан на сугубо акустическом воздействии на тело человека.]
К настоящему времени известны многочисленные механизмы управления работой генома и процессами развития, имеющие сугубо химическую природу. В последующем изложении мы не будем упоминать их потому, что они уже были объектами многих исследований, и не составляют предмета нашего рассмотрения. Но молчание по их поводу не нужно расценивать как свидетельство игнорирования этих механизмов. Напротив, мы считаем их важными компонентами информационной системы организма. Просто в проблеме формирования многоклеточного организма не они оказались главными.
Главное средство управления биохимическими процессами в клетках — локальное разрыхление хроматина — никак не удается свести к некому химическому агенту, проявляющему химические предпочтения. Оно способно затронуть любой (в химическом смысле) ген, но почему-то, в большинстве случаев, зависит от расположения клетки в организме, что совершенно нехарактерно для химических процессов. Это ярко обнаруживается при разрыхлении гена внутри сплошной массы неактивного гетерохроматина, содержащего разные гены, в том числе и точно такие же как данный.
Подобные факты говорят в пользу физической природы процесса разрыхления, безразличного к химическим особенностям генов, но учитывающего их координаты.
Ряд данных говорит не только в пользу волнового механизма разрыхления хроматина, но и в пользу определенной пространственной организации волн, например, фокусировки. Именно действие сфокусированных колебаний способно объяснить выборочное разрыхление хроматина в компактной зоне, окруженной массивом плотного гетерохроматина. В пользу пространственной организации волновых процессов, управляющих генами, говорят и закономерности пространственного расположения генов эукариот, и их пространственная фиксация.
При молекулярных размерах генов избирательное волновое управление их разрыхлением должно представлять собой непростую задачу. Трудно создать интенсивные колебания в малой зоне, не затронув соседние зоны, где расположены другие гены, не требующие активирования. Прокариотам, использующим химические, контактные способы активирования генов, легче обеспечить избирательное управление транскрипцией. Их гены могут располагаться вплотную друг к другу без опасений непроизвольного активирования соседнего гена.
Есть ли свидетельства того, что Природе было труднее обеспечить раздельное, независимое активирование генов эукариот по сравнению с прокариотами? Да, можно назвать, по крайней мере, два таких факта.
1.Во-первых, гены эукариот, в отличие от прокариот, не расположены вплотную друг к другу — между ними вставлены неинформативные спейсеры (разделители, от англ. spacer — прокладка, распорка), вдесятеро и более превышающие по длине сами гены и соответственно увеличивающие общую массу генома.
2.Во-вторых, точность управления активированием генов у эукариот значительно снижена. В отличие от прокариот, у них, хотя и с малой скоростью, синтезируются РНК почти со всех имеющихся генов.
По мнению исследователей, «...можно предположить, что слабая функция генов ... отражает ... несовершенство регуляторного аппарата, запирающего гены. Что-то вроде неплотно прикрытых «подтекающих» кранов» [Нейфах, Лозовская, 1984]. Так что признаки волнового механизма активирования генов закономерно проявились и в этом.
Характерно то, что считывание информации с ДНК происходит не только при транскрипции, но и при репликации — т.е. при удвоении числа хромосом перед делением клетки. С химической точки зрения эти процессы очень похожи друг на друга — в одном случае на матрице ДНК синтезируется нить РНК, а в другом — нить ДНК. В половине случаев (когда матрицей служит так называемая «отстающая» нить ДНК) процесс начинается совершенно одинаково — синтезируется нить РНК. В случае репликации, образовавшийся праймер (т.е. «затравка») в дальнейшем замещается цепочкой ДНК, а с 11-го нуклеотида и сам синтез цепочки нуклеотидов переключается на производство ДНК. Но, несмотря на такое сходство, репликация не зависит от разрыхления хроматина, от связи ДНК с нуклеосомами, а транскрипция — зависит.
Если для синтеза первого десятка нуклеотидов РНК не нужен отрыв ДНК от нуклеосом, то почему такой отрыв становится необходимым, когда дело доходит до синтеза РНК, несущих коды белков, причем это наблюдается у всех эукариот и, следовательно, жестко контролируется естественным отбором?
С химической точки зрения, отличие продуктов репликации (ДНК) от продуктов транскрипции (РНК) — в том, что в сахаре, содержащемся в цепочке нуклеотидов, одна гидроксильная группа заменена атомом водорода, а урацил (один из четырех нуклеотидов РНК) заменен тимином, где место одного из атомов водорода заняла метильная группа. В общей сложной структуре цепочки нуклеотидов эти различия незначительны и сами по себе не могут ответить на сакраментальный вопрос — почему, в итоге, для получения молекул РНК требуется разрыхление хроматина, а для получения ДНК оно не требуется?
Только КСГ разъясняет эту странную ситуацию. Формирование многоклеточных организмов требует учета расположения клеток в трехмерной структуре, и для управления таким процессом Природа использовала принцип оптической проекции активных зон волновых полей организма на гены. Механизм разрыхления хроматина выбран Природой как способ управления активностью генов со стороны волнового поля. Этот же способ управления обусловил удивительную консервативность аминокислотных последовательностей некоторых гистонов, без чего не было бы стабильности порогового усилия, при котором нуклеосомы начинают отрываться от ДНК.
Управление активированием генов у всех эукариот через разрыхление хроматина показывает, что, при всем многообразии химических механизмов управления внутриклеточными процессами, именно волновой механизм управления транскрипцией оказался для эукариот решающим.
Вот, почему ДНК эукариот — в отличие от прокариот — «намотана» на нуклеосомы и, соответственно, изменены ферменты, синтезирующие у эукариот новую ДНК (ДНК-полимераза) и РНК (РНК-полимеразы). Первый фермент приобрел способность работать независимо от разрыхления хроматина (такой проблемы не было у прокариот), а второй (точнее — РНК-полимеразы I и II) — только при разрыхленном хроматине. Те загадки, о которых мы говорили выше, появились как результат выработанных естественным отбором особенностей ДНК- и РНК-полимераз эукариот.
Чтобы оценить степень соответствия между следствиями КСГ и биологическими фактами, удобно одновременно сравнивать факты со следствиями теории диссипативных структур (ТДС). Поэтому отметим, что ТДС не объясняет необходимости разрыхления хроматина для синтеза РНК, притом, что синтез ДНК происходит без такого разрыхления. А для КСГ этот же факт стал одной из фундаментальных опор.
F138
--------------------------------------------------------------------------------
Исследуя механизмы шока, Ревич обнаружил потрясающий эффект: если подопытное животное ввергается в состояние сверхострого шока — наступает распространенное «продырявливание» многих частей тела.
Небольшое пояснение. Сверхострый шок у животных легко вызывается внутривенным введением коллоидного металла (см. ниже), ошпариванием горячей водой или путем нанесения тяжелой механической травмы. Для следующего вслед за этим состояния наиболее характерными и очевидными являются страшные симптомы со стороны центральной нервной системы, включая экзофтальм (выпучивание глаз) и паралич задних конечностей, за которым следуют судорожные движения, обычно с летальным исходом. Точно такой же сверхострый шок наблюдается у людей после переливания крови, несовместимой по групповому признаку, или после очень тяжелой травмы.
Чтобы детально изучить изменения, происходящие в организме вследствие сверхострого шока, Ревич «по пояс» макал подопытных крыс в кипяток, после чего извлекал из них разные части тела и проводил гистологический анализ. Так вот самым характерным нарушением при сверхостром шоке оказалась клеточная вакуолизация, т.е. появление «дырки» внутри изначально целостной клетки. Такие вакуоли присутствуют в клетках паренхимы печени, в меньшей степени — в альвеолярных клетках легкого, и еще в меньшем количестве — в клетках почек. Но особый интерес вызывает обнаружение вакуолей в цитоплазме и даже в ядрах клеток головного мозга — именно этот факт объясняет манифестацию при сверхостром шоке тяжелых неврологических симптомов.
При менее тяжелых формах шока вакуолизация клеток также имеет место быть, но в существенно меньшем объеме — главные патологические изменения затрагивают мышцы. Кроме всего прочего, эти изменения проявляются сосудистой и интерстициальной патологией, такой как отеки и капиллярные кровотечения. Особыми характеристиками состояния обычного (не сверхострого и не острого) шока являются два других специфических патологических проявления: а) милиарные поражения слизистой оболочки желудка, ведущие к кровотечению и изъязвлению, и б) явное накопление жидкости в первой части тонкого кишечника.
Возникает закономерный вопрос:
Каким образом в изначально целостных клетках без какого либо нарушения их оболочки могут внезапно массовым порядком возникать полости?
Разве не линзоподобной фокусировкой химических волн на клеточных оболочках с концентрацией преломленного энергетического пучка внутри клетки, происходящей в экстремальных обстоятельствах, вызываются эти чрезвычайно странные явления? Кстати, если положить сырое яйцо в микроволновую печь, то что будет?
Изучая изменения, происходящие в клетках вследствие облучения, Ревич обнаружил, что после облучения — особенно при использовании высоких доз — также наблюдается интенсивная клеточная вакуолизация. С помощью радиоактивного натрия было также установлено, что после облучения наблюдается большая склонность этого, по большей части внеклеточного элемента проникать в клетки.
Вакуоли Но дальше все оказалось еще интереснее — введение в организм конъюгированных жирных кислот приводило к появлению вакуолей не только в цитоплазме, но даже в ядрах клеток! Иными словами, воздействие на клетки конъюгированных жирных кислот практически повторяет таковое радиации.
Но дальше все оказалось еще интереснее — введение в организм конъюгированных жирных кислот приводило к появлению вакуолей не только в цитоплазме, но даже в ядрах клеток! Иными словами, воздействие на клетки конъюгированных жирных кислот практически повторяет таковое радиации.
Эффект от препаратов Ревича на основе селена (Se) на клеточном уровне виден даже при использовании дозировки в микрограммах — в цитоплазме клеток происходит вакуолизация! Интересно также отметить в этой связи, что, несмотря на вакуолизацию клеток, наблюдаются также и околоклеточные отеки (рис. 1).
Вакуоли в эритроцитах Обработка in vitro эритроцитов конъюгированными жирными кислотами — особенно триенами — вызывает выраженную, хорошо заметную вакуолизацию (рис. 2).
А вот в присутствии антагонистов жирных кислот, к которым относятся некоторые спирты, во многих случаях предотвращаются и вакуолизация, и даже изменения в ядрах, наблюдающиеся у облученных животных контрольной группы. Кстати сказать, кому неизвестно хоть и слабое, но все же реально существующее противолучевое действие этилового спирта?!
Стоит также упомянуть и о двух других, чрезвычайно интересных опытах Ревича.
1.Были приготовлены 1/10-молярные растворы с рН=8.5 двухосновных фосфатов лития (Li), натрия (Na), калия (K), аммония (NH4) и рубидия (Rb). Каждый из растворов вводили внутривенно мышам по ¼ мл в минуту — вплоть до момента смерти животного. После этого их органы — особенно головной мозг — немедленно фиксировались в жидкости Буэна и изучались гистологически. В случаях аммония и рубидия наблюдали присутствие вакуолей в клетках и, особенно, в ядрах! При введении калия вакуоли появлялись в цитоплазме, а после применения натрия отмечен только лишь околоклеточный отек. При введении лития никаких изменений не отмечено.
2.Лейкоциты с колларголомНормальные лейкоциты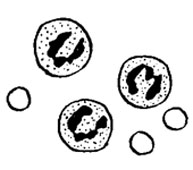 В экссудат, полученный путем плевральной пункции и богатый гранулоцитами, добавляли небольшое количество коллоидного серебро-белкового препарата (0.1% Колларгола) и содержали при температуре +38°C. Было установлено, что, вслед за фагоцитозом крупинок серебра, это вызывало появление вакуолей в лейкоцитах. Причем вакуоли быстро увеличивались в размерах с последующим взрывом лейкоцитов! Ядра же оставались в виде ядерных «теней». На рисунке показаны нормальные гранулоциты и, рядом, вакуолизированные гранулоциты на грани взрыва с поглощенными крупинками колларгола.
В экссудат, полученный путем плевральной пункции и богатый гранулоцитами, добавляли небольшое количество коллоидного серебро-белкового препарата (0.1% Колларгола) и содержали при температуре +38°C. Было установлено, что, вслед за фагоцитозом крупинок серебра, это вызывало появление вакуолей в лейкоцитах. Причем вакуоли быстро увеличивались в размерах с последующим взрывом лейкоцитов! Ядра же оставались в виде ядерных «теней». На рисунке показаны нормальные гранулоциты и, рядом, вакуолизированные гранулоциты на грани взрыва с поглощенными крупинками колларгола. 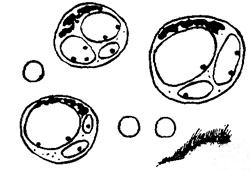
А теперь самое интересное: добавление минимальных количеств морфина или иных дериватов опия полностью предотвращает эти изменения в лейкоцитах — вакуоли не обнаруживаются и лейкоциты не взрываются. Причем оказалось, что способностью предотвращать взрыв лейкоцитов обладают только морфин и прочие опиумные алкалоиды. Ни адреналин, ни хинин, ни прочие исследованные вещества никакого эффекта не оказывают.
www.ling1.net/
читать дальшеF134
--------------------------------------------------------------------------------
В генетике многоклеточных организмов нельзя пройти мимо загадки целостности организма. Вероятно, нужно искать не способ кодирования каких-то дискретностей, а способ записи информации о неразрывной трехмерной (а с учетом времени — четырехмерной) анатомии всего организма.
«Из открытий Менделя, апробированных победным развитием генетики ... вытекает бесспорный принцип дискретности наследственной детерминации признаков. Однако бесспорно и то, что эмбриология не имеет оснований говорить о независимом возникновении и развитии признаков в онтогенезе. Нормальный онтогенез — это совокупность процессов изменения состояния целостности. Что означает противоречивость двух бесспорных утверждений?» [Токин, 1987]
Загадка целостности организма в индивидуальном развитии многократно приводила исследователей к идее биополя, управляющего пространственной организацией живой материи. Иногда авторы не говорили о биополе напрямую, но логикой фактов подводили к мысли о его существовании.
Термин «биополе» используется не только в биологии развития, где его применяют к внутриорганизменным процессам полевой природы. Термин «биополе» используется также биофизиками, экстрасенсами и др., но уже в ином смысле — применительно к процессам полевой природы в пространстве, окружающем организм. Мы будем рассматривать только процессы, протекающие внутри организмов.
Как отмечалось ранее, иногда формы одного животного (контур рыбы, кость птицы и т.п.) удается точно совместить с формами животного другого биологического вида, если подвергнуть изображение простым деформациям — плавному растяжению, сжатию или др. [Thomрson, 1942]. Здесь тоже видны явления полевой природы, проглядывают законы, которые нельзя выразить в одномерном или даже в двумерном пространстве. Глазам исследователей предстает явная передача от предков к далеким потомкам не только биохимических процессов, но и пространственного образа, способного плавно деформироваться в ходе эволюции. Формирование пространственного образа подразумевает протекание неких пространственно-организованных событий, наводя на мысль о биополе.
Обстоятельный (хотя далеко не полный) обзор разработок проблемы биополя приведен в работе Токина от 1987 года.
Довольно развитые представления о биополе изложил П. Вейс, уподобивший его системе векторов. Представления Вейса явились результатом его исследований по регенерации органов. По Вейсу, поле материально, но его природа не может быть сведена к химической, так как, по его мнению, химические факторы способны лишь активировать, но не формировать поле.
Н. К. Кольцов на основе изучения процессов в ооцитах и яйцах создал представление об управляющем воздействии на зародыш ряда «силовых» полей в виде разностей электрических, химических, температурных, гравитационных, диффузионных и других потенциалов. В ходе развития силовое поле зародыша усложняется, дифференцируется, но остается единым.
Близкую к взглядам Н. К. Кольцова трактовку морфогенетических процессов предложил в 1958 г. Б. Вейсберг, который на основе изучения колебаний электрических потенциалов у миксомицетов создал представление о «колебательных полях».
Идейную близость к подобным взглядам можно заметить в теории «аксиальных градиен-тов» американского биолога Ч. Чайлда.
Интересные этапы проходила концепция биополя, разрабатывавшаяся Александром Гавриловичем Гурвичем [Гурвич, 1944; Любищев, Гурвич, 1998]. Отметим некоторые из его тезисов, хотя подвергнутые критике, но, как мы увидим далее, хорошо отражающие реальность:
•поведение элементов всех микроморфогенетических процессов управляется единым фактором — «преформированной морфой», задающей еще не существующую структуру и определяющей окончательную конфигурацию зачатка;
•каждая клетка имеет собственное поле, действие которого выходит за пределы клетки; при делении клетки делится и ее поле; клетки оказывают своими полями влияние друг на друга; по мере удаления влияние поля быстро ослабевает;
•в ходе индивидуального развития происходит эволюция «поля целого»; то целое, что существует в данный момент, определяет ход дальнейшего развития.
Даже краткий обзор полевых концепций в биологии и связанных с ними исследований занял бы много места. Идею существования в организме некого биополя, участвующего в управлении морфогенезом, в разное время и с разных позиций развивали, например, И. А. Аршавский, Л. В. Белоусов, Г. Р. де Бер, Т. Бовери, Б. Брайент, Л. Вольперт, Дж. С. Гексли, Е. Гиено, В. В. Исаева, Г. П. Короткова, А. А. Любищев, В. М. Маресин, Е. В. Преснов, П. Г. Светлов, К. Уоддингтон, П. Френч, Д. С. Чернавский, Г. Шпеман, Т. Ямада и другие ученые. Их взгляды базировались на глубоких исследованиях, оставивших заметный след в науке.
К сожалению, сильнее оказалось отрицательное влияние многочисленных энтузиастов, создавших вокруг проблемы ажиотаж, но не приблизивших понимание природы явления. Идея биополя оказалась серьезно дискредитированной ими, так что сегодня биологи упоминают ее преимущественно в уничижительном тоне.
Сегодня ясно, что исследования нескольких десятилетий принципиально не могли раскрыть природу внутреннего биополя организма, поскольку этому должно было предшествовать фундаментальное открытие в иной области. Но изложим события по порядку.
Идея биополя близка к идее волнового информационного взаимодействия организма с генами. Мысль о том, что формирование многоклеточного организма связано с передачей информации волновыми полями, имеет немалую предысторию, она высказывалась рядом исследователей. Например, А. Н. Мосолов [1980] предположил, что на гены воздействует акустическое поле. Но акустические (и электромагнитные) колебания, которые могли бы взаимодействовать с генами (т.е. имели бы протяженность фронта волны, сопоставимую с размером гена) — такие колебания очень сильно затухают в биологических тканях, отчего бессмысленно говорить об их распространении более чем на миллиметры. В то же время, чтобы связать формирование организма с волновым управлением генами, нужны волны, способные без чрезмерного затухания распространяться по всему организму.
Ситуация казалась безвыходной, пока не были открыты волны совершенно новой природы. Стремясь создать удобную для экспериментов упрощенную модель цикла Кребса — цепочки реакций, питающих наш организм энергией, Борис Павлович Белоусов открыл в 1951 году колебательную химическую реакцию. Наблюдать такую реакцию очень легко; достаточно смешать в пробирке 10мл водного раствора веществ:
•лимонная кислота — 2,00 г
•сульфат церия — 0,16 г
•бромат калия — 0,20 г
•серная кислота (1:3) — 2,0 мл
•воды до общего объема — 10,0 мл.
При комнатной температуре в пробирке возникают периодические изменения цвета «желтый-бесцветный» с периодом порядка десятков секунд. Более контрастное изменение цвета «синий-красный» происходит при добавлении к смеси железофенантролина в качестве индикатора. Нагревание ускоряет смену цветов. Постепенное расходование ингредиентов ведет к затуханию процесса, а прибавление израсходованного вещества возобновляет его.
Однако химические журналы отказались публиковать сообщение о колебательной реакции, считая статью блефом. Лишь в 1959 г. появилось короткое сообщение в журнале по радиационной медицине, который химики не читают. В 1970 г. Б. П. Белоусов умер, так и не сумев донести открытие до научной общественности . А в 1980 году за это открытие была присуждена Государственная премия. С легкой руки Ильи Романовича Пригожина — президента Бельгийской Королевской академии наук — открытие Б. П. Белоусова было названо самым выдающимся экспериментом ХХ века.
Реакция БЖ
К этому времени А. М. Жаботинский и А. Н. Заикин установили, что колебательные реакции способны распространяться в соответствующей среде в виде волн. На фото запечатлен один из моментов распространения волн химических реакций в тонком слое жидкости, налитой на дно плоского сосуда.Такие волны назвали волнами Белоусова-Жаботинского (БЖ). Они, как лесной пожар, черпают энергию из среды, по которой распространяются, отчего, даже при очень серьезных потерях энергии, способны распространяться на неограниченно большие расстояния.
Открытие колебательных реакций, как часто бывает, стало результатом удачного стечения обстоятельств. Если бы период изменения цвета в пробирке измерялся сотыми долями секунды или, наоборот, многими часами и днями, то явление осталось бы незамеченным. Между тем, есть основания считать, что подобные реакции с сильно отличающимися временными характеристиками реально существуют и достаточно распространены. В этом смысле привлекает внимание причудливый узор разрезанного агата. Похоже, что он изображает застывшие волны, связанные с химическими процессами, но протекавшие во много раз медленнее, чем в случае волн БЖ.
Среди многообразия химических реакций есть и такие, что распространяются с очень высокими скоростями, например, взрывы, детонация в двигателях и др. Для скорости продвижения химических волн важен механизм их распространения. Если в его основе лежит диффузия молекул, то скорость волны будет зависеть от размера молекул, температуры и вязкости среды. Однако агентами, инициирующими реакцию в соседних зонах, могут быть также электроны, протоны, кванты излучения, рождающиеся в ходе реакции, и они определят другие, гораздо более высокие скорости ее распространения.
Это дает основания предполагать, что среди большого числа реакций, протекающих в протоплазме клеток, существуют и такие, что распространяются в виде волн, хотя они, из-за временных характеристик или по другим причинам, пока напрямую не обнаружены. Неслучайно Б. П. Белоусов открыл колебательные реакции, пытаясь моделировать едва ли не самую распространенную в живой природе цепочку биохимических процессов — цикл Кребса.
Один из исследователей [Pohl, 1983] на основании двух независимых экспериментальных методик выяснил, что, по-видимому, волны химических реакций возникают в протоплазме каждой нормальной эукариотической клетки. Тем самым, подкрепилось предположение, что волны, переносящие информацию при формировании многоклеточного организма, являются волнами химических реакций.
Веские основания искать в организме химические колебательные реакции дает картина переменной электрической активности тканей, которую нельзя свести только к активности нейронов и мышц. Например, описано появление особой компоненты колебаний электрического потенциала во время восстановления (регенерации) сломанной кости лапы кролика [Бутуханов, Ипполитова, 1982].
F135
--------------------------------------------------------------------------------
Волны химических реакций, среди которых особую известность получили волны БЖ, оказались неожиданными не только по своей природе. Как явствовало уже из первых осциллограмм, полученных Б. П. Белоусовым, они относились к особому классу — к классу уединенных волн или солитонов.
В настоящее время студенты ВУЗов изучают волновые процессы на примере синусоидальных колебаний, и лишь немногие из них получают теоретические представления о солитонах, которые являются единственным типом волн физиологических процессов. Уже изучая практические предметы, биологи и медики, не зная теории, сталкиваются с солитонами в виде нервных импульсов, волн сокращений миокарда, разрядов электрического ската, волн перистальтики, волн нервного возбуждения в тканях мозга и т.д. Поэтому есть смысл сказать несколько слов о солитонах вообще.
Физики и математики делят колебания на линейные и нелинейные. Теория линейных колебаний создана в XVIII-XIX веках и лежит в основе аппарата современной математической физики. Синусоидальные колебания — это линейные колебания. Систематическая теория нелинейных колебаний и волн — теория солитонов — появилась во второй половине XX века. Познакомиться с ней можно по работам [Филиппов, 1986; Додд и др., 1988; Ньюэлл, 1989] и многим другим.
Синусоидальные колебания в чистом виде — наиболее простые колебания, колебания одной определенной частоты, строго симметричные относительно исходного уровня. Для прохождения синусоидального колебания по каналам связи требуется минимальная ширина полосы частот. Все это обусловило широкое использование синусоидальных колебаний в технике, их почетное место в учебных программах.
Солитоны, как правило, распространяются в нелинейных средах, что существенно влияет на их особенности.
1.Во-первых, нелинейность среды определяет некоторую оптимальную амплитуду солитона, от которой ему, в отличие от синусоидальных колебаний, труднее отклоняться и в меньшую, и в большую стороны.
2.Во-вторых, нелинейность среды создает для него некую возможность обмена одних параметров на другие, скажем, амплитуды на форму. Последнее придает солитону способность распространяться без заметных изменений амплитуды на гораздо большие расстояния, чем мы ожидали бы по опыту работы с синусоидальными волнами.
3.Наконец, в-третьих, нельзя не отметить, что нелинейность среды распространения солитонов часто определяется активностью среды и это особенно характерно для биологических тканей. В таких случаях размах и форма проходящего солитона имеют повышенную стабильность, потому что по мере движения солитона среда подпитывает его своей энергией.
Пожалуй, наиболее существенным свойством солитонов, резко отличающим их от синусоидальных колебаний, является независимость друг от друга. У синусоиды нельзя отделить одну волну от других или сдвинуть относительно других, изменить размах. Это сразу превратит синусоиду определенной частоты в нечто иное — в сложную совокупность разночастотных синусоидальных колебаний. Ничего похожего не грозит солитонам. Каждый из них можно, например, смещать во времени, и это не изменит ни его основных параметров, ни параметров других солитонов.
Соответственно, по отношению к солитонам нужно с большой осторожностью говорить о частоте. Они могут следовать на одинаковом расстоянии друг от друга (тогда обретает смысл понятие частоты следования), но могут от волны к волне резко изменять интервал или вовсе следовать через хаотически меняющиеся промежутки времени. В биологических процессах встречаются все эти случаи.
Солитон
Типичная форма солитона показана на рисунке справа. Она может существенно изменяться в зависимости от природы процесса и параметров среды. Например, затухающие колебания за задним фронтом волны возникают только при существенном ухудшении условий прохождения высоких частот.Часто солитоны обладают большой скважностью, т.е. расстояние (во времени) между соседними волнами оказывается во много раз большим, чем длительность самого колебания. В таких случаях энергия колебаний сосредотачивается на гораздо более высоких частотах, чем частота следования волн.
При резком различии скоростей процессов, определяющих передний и задний фронты волны, задний фронт может превратиться в медленно спадающую кривую, что придает солитону вид одиночного скачка параметра с последующим длительным (по сравнению с продолжительностью переднего фронта) возвращением в исходное состояние. Такую разновидность солитонов иногда называют кинками. Волны Белоусова-Жаботинского имеют именно такую форму — у них процессы, определяющие задний фронт волны, протекают намного медленнее, чем процессы, от которых зависит крутизна переднего фронта.
Способность солитонов следовать друг за другом через произвольные интервалы времени открывает для них большие возможности в части переноса информации. Кроме того, солитон, по сравнению с синусоидальными колебаниями, очень устойчив против возмущений, вносимых неоднородностью среды распространения и внешними помехами [Захаров, Шабат, 1962]. Эти свойства приобретают особую значимость при переносе информации в такой неоднородной и слабо защищенной от внешних воздействий среде, как живой организм.
Многие математики не согласны называть волны химических реакций солитонами. По их мнению, химические волны и солитоны — это разные понятия, разные категории объектов. Но в таком возражении проявляется лишь желание математиков подогнать понятие солитона под класс объектов, которые они научились описывать. Волны химических реакций они пока описывать не умеют. Между тем, многие специалисты вполне логично рассматривают солитон как нелинейное устойчивое локализованное возбуждение (например, [Шахбазов и др., 1996]), а такая трактовка охватывает и волны химических реакций.
Оптика нового типа
«Возможно такое, что вам и не снилось... Я буду рассказывать, как устроена Природа, и, если вам не понравится, как она устроена, это будет мешать вашему пониманию. Физики научились решать эту проблему: они поняли, что, нравится им теория или нет — неважно. Важно другое — дает ли теория предсказания, которые согласуются с экспериментом. Тут не имеет значения, хороша ли теория с философской точки зрения, легка ли для понимания, безупречна ли с точки зрения здравого смысла. Квантовая электродинамика дает совершенно абсурдное с точки зрения здравого смысла описание Природы. Но оно полностью соответствует эксперименту. Так что я надеюсь, что вы сможете принять Природу такой, какая она есть — абсурдной».
Фейнман, 1988
Сам факт осциллографической регистрации Б. П. Белоусовым химических колебаний показал, что в ходе реакции скачкообразно изменяется окислительно-восстановительный потенциал среды. Как оказалось, химической волне сопутствует скачок электрического потенциала порядка десятых или сотых долей вольта. Но есть ли основания считать, что подобные волны возникают в живых организмах?
Непосредственная регистрация в организме колебаний с ожидаемыми параметрами встречает серьезные технические трудности. Косвенные данные говорят, что длительность переднего фронта ожидаемых волн имеет временную протяженность порядка десятков пикосекунд. Это значит, что основная энергия колебаний сосредоточена в области десятков гигагерц. К тому же, мощность колебаний в расчете на одну клетку очень мала. Самые современные осциллографы еще не способны зарегистрировать подобные волны, показать их форму.
Учитывая, что увидеть осциллограмму структурогенных волн пока не удается, Герберт Пол заменил прямую регистрацию косвенными исследованиями на основе микродиэлектрофореза. В раствор, окружающий выделенную живую клетку, Пол ввел мельчайшие гранулы диэлектрика и наблюдал под микроскопом их поведение. В зависимости от того, у какого вещества — у жидкости или у гранул — была выше диэлектрическая проницаемость, частицы двигались к клетке, либо в противоположную сторону [Pohl, 1983]. Это четко указывало на существование в клетке переменного электрического поля. Но оставалось неясным, привязаны ли электрические колебания к поверхности клеток или же они протекают в объеме цитоплазмы?
На этот вопрос ответила другая серия опытов. Пол создал вокруг взвешенной в жидкости клетки вращающееся электрическое поле, и увидел, что клетка тоже стала вращаться, хотя и с меньшей скоростью. Более того, при изменении скорости вращения поля клетка обнаружила резонансные свойства — на определенных частотах она резко ускоряла свое вращение. Этот эффект, названный клеточным спиновым резонансом, противоречил предположению об электрических колебаниях на так называемой мембране, но говорил в пользу химических волновых процессов в протоплазме клетки.
Что произойдет при движении волны химических реакций в протоплазме клетки, когда фронт волны пересечет клеточное ядро?
В кольцевой зоне оболочки клеточного ядра, где в какой-то момент расположен скачок окислительно-восстановительного потенциала, возникнет очень высокая напряженность электрического поля. Скачок потенциала порядка десятых долей вольта воздействует на поверхностный слой (ошибочно называемый мембраной) общей толщиной всего 14 нм и создает в нем напряженность поля до 200 кВ/см. Вследствие этого в зоне соприкосновения фронта волны с ядерной оболочкой должно произойти интенсивное электрострикционное сжатие диэлектрического материала оболочки. Согласно расчетам [Артюшин, Барбараш, 1985], если бы отсутствовало гидравлическое сопротивление внутриядерной жидкости, то деформация оболочки достигла бы 10% диаметра ядра.
Быстрое перемещение кольцевой зоны деформаций по оболочке ядра порождает во внутриядерной жидкости акустические колебания. Скорости волн различной природы обычно не совпадают. Поэтому на границе раздела — на оболочке ядра, где энергия химических волн переходит в энергию акустических, происходит преломление волнового поля. Оболочка ядра является, в первом приближении, сферой. Явления, связанные с преломлением волнового поля на сферической поверхности, досконально изучены оптиками — такая поверхность приобретает свойства линзы. Конечно, того, что принято называть линзой, здесь нет. Но это не меняет результата. Возможно, следует ввести новый термин — линза-мембрана, мембролинза или что-либо в этом роде.
В обычной оптике преломляющиеся волны не изменяют свою природу в момент преломления, т.е. при переходе из области с одной скоростью распространения в область с другой скоростью. В обычной оптике изменение скорости распространения объясняется не изменением природы волнового поля, а изменением свойств среды распространения. Такую оптику можно условно назвать гомоволновой (homos — одинаковый). В нашем же случае изменение скорости вызывается именно изменением природы волнового поля. Здесь принципиально важно, что волны одной природы при достижении некой преобразующей пленки превращаются (передают свою энергию) в волны иной природы. Оптику этого неизвестного ранее типа можно назвать гетероволновой оптикой (heteros — иной, другой).
Гетероволновая оптика обладает рядом особенностей. Поскольку в ней могут сочетаться волновые процессы с сильно различающимися скоростями распространения, то оказываются достижимыми очень высокие коэффициенты преломления, а это позволяет при прочих равных условиях резко сократить количество компонентов оптической схемы и уменьшить аберрации. Если пленка-преобразователь работает на основе необратимого физического явления (например, в оболочке клеточного ядра возникает необратимая электрострикция, а не обратный пьезоэлектрический эффект), то из высокого коэффициента преломления не следуют, как это обычно бывает, высокие потери на отражение (так как не индуцируется противоволна). В гетероволновой оптике фокусирующими элементами служат не линзы, а тонкие пленки, из-за чего такая оптика имеет очень низкую материалоемкость.
Таким образом, оболочка ядра, действуя как линза гетероволновой оптики, преобразует химические волны протоплазмы в акустические колебания внутриядерной жидкости.
Термин «оптика» имеет два толкования — узкое и расширенное. В узком понимании это наука о том, что происходит с лучами видимой части спектра электромагнитных колебаний при их отражении, преломлении, поглощении и т.д. В расширенном понимании оптика охватывает процессы, в которых могут участвовать не только электромагнитные волны видимой части спектра, но и другие волны и объекты, поведение которых описывается сходными законами.
В расширенном понимании для разных конкретных задач термин «оптика» обычно дополняется уточняющим словом — «инфракрасная оптика», «рентгеновская оптика», «акустическая оптика», «электронная оптика», «оптика протонов» и т.д. Взаимодействие химических волн протоплазмы с клеточным ядром тоже описывается законами оптики, что дает основания отнести его к оптике в расширенном понимании. Соответственно, и здесь термин требует уточнения. Поскольку процессы связаны с ядром клетки, данный раздел оптики уместно назвать кариооптикой или сокращенно — К-оптикой.
Кариооптика
Представим себе, как показано на рисунке, что точка A относится к химическому волновому полю организма, т.е. находится вне ядра, а точка C является проекцией точки A в акустическом поле ядра. Эти точки расположатся на прямой, проходящей через центр ядра, и будут разделены ядерной оболочкой.Обозначим: R — радиус ядра; x — расстояние между точкой A и ближайшей точкой В на поверхности ядра; y — расстояние от точки В до точки фокусировки С; n — отношение скоростей распространения химических и акустических волн (коэффициент преломления).
Тогда, на основании формулы нулевого инварианта Аббе [Ландсберг, 1976] получим:
nRx
y= —————— ( т.е. у = nRx/(n-1)x + R )
(n-1)x + R
Анализ этой формулы, с учетом реальных размеров клеток животных и их ядер, показывает, что при n ≥ 2.7, все пространство организма, как бы велик он ни был, проецируется внутрь ядра каждой клетки [Артюшин, Барбараш, 1985]. Такой характер проекции не нарушается при изменениях размеров ядер.
Здесь мы столкнулись с явлениями, относящимися к химии, физике диэлектриков, теории нелинейных колебаний, оптике, биологии, а вовсе не к медицине. Именно поэтому до тех пор, пока проблемами здоровья человека будут заниматься именно медики, эту область естествознания будет окутывать полный мрак.
F136
--------------------------------------------------------------------------------
«Продвижение науки вперед часто происходит тогда, когда перед нами раскрывается неведомая доселе сторона вещей, что обусловлено не столько применением какого-то нового метода, сколько рассмотрением объектов под другим углом зрения».
©Jacob, 1977
На основе открытия колебательных реакций и волн Белоусова-Жаботинского, еще в 1982 году была разработана концепция структурогенеза (в дальнейшем — КСГ), включившая в себя впервые сформулированную идею гетероволновой оптики и объяснившая на такой основе принципы генетического кодирования структуры многоклеточного организма.
Очень важной для этой работы была исходная мысль Б. П. Белоусова о сходстве волновых химических реакций с процессами в живом организме. Такой подход представил известные биологические факты в новом свете — явления и закономерности, ранее казавшиеся несвязанными между собой, выстроились в четкую систему.
Согласно КСГ, в протоплазме клеток многоклеточного организма периодически самовозбуждаются волны химических реакций, пробегающие по всему организму и потому несущие информацию о его сиюминутном строении.
В протоплазме клеток бодрствующего животного такие волны маломощны и относительно разрозненны — по степени упорядоченности их можно сравнить с мелкой рябью на поверхности воды. Роль таких волн ограничивается передачей информации на малые расстояния — порядка десятков клеток.
Однако, кроме того, в определенной фазе сна, при заторможенности скелетных мышц, в паузах ритмики сердца и легких, кратковременно возникают мощные пакеты химических волн такой же природы, охватывающие весь организм. При всем их сходстве с волнами БЖ, эти волны нельзя отождествлять — у них разный химизм, разные механизмы распространения и сильно отличающиеся количественные характеристики.
Волны БЖ распространяются в растворе благодаря диффузии молекул, что определяет невысокие скорости продвижения волны — порядка миллиметров в секунду. Но диффузия активных молекул — не единственный фактор, способный инициировать реакцию в растворе. Хотя химизм структурогенных волн пока непонятен (вспомним, что от открытия Г. Менделем генов до выяснения их химической природы прошло около 90 лет!), есть веские основания предполагать, что продвижение фронта структурогенной волны обеспечивают не диффундирующие молекулы, а более быстрые частицы, обладающие, к тому же, очень коротким пробегом.
Это заставляет вспомнить об открытом А. Г. Гурвичем слабом ультрафиолетовом излучении живых клеток, которое он назвал митогенетическим. Название точно отразило подмеченную ученым связь интенсивности излучения с делением клеток (митозами), т.е. с процессами развития. Излучение универсально по отношению к живым клеткам разных типов — оно усиливается при стрессовом состоянии клетки и постепенно затухает после ее гибели. Все это заставляет думать, что УФ-кванты порождаются биохимическими реакциями, в норме свойственными клетке.
Обладая скоростью света, УФ-кванты должны проходить расстояние между последовательно вступающими в реакцию молекулами намного быстрее чем диффундирующие атомы, молекулы или радикалы. Если химическая реакция, генерирующая митогенетическое излучение, проходит по раствору в виде волны, то от такой волны нужно ожидать гораздо более высокой скорости распространения по сравнению с уже знакомыми нам волнами БЖ. Вероятно, основные потери времени приходятся не на перемещение УФ-квантов в пространстве, а на их реакцию с субстратом, т.е. на паузу между поглощением молекулой кванта и генерированием новой порции квантов.
Выше было отмечено, что проекция многоклеточного организма на хроматин ядра имеет место при коэффициенте преломления порядка 2,7 и более. Скорость акустических волн внутри клеточного ядра близка к скорости звука в воде, т.е. около 1,5 км/с. Следовательно, нужно ожидать скорости распространения химических волн порядка 1,5 х 2,7 = 4,05 км/с и выше.
Такая скорость распространения структурогенных химических волн на несколько порядков выше скорости известных волн БЖ, что как раз и говорит в пользу распространения волн в протоплазме не за счет диффузии «горячих» молекул, а благодаря упомянутым квантам ультрафиолетового излучения.
У прокариот, использующих чисто химические способы управления активностью генов, включение и выключение транскрипции осуществляют специфичные регуляторные белки. Особенности геномов эукариот заставляют думать, что их способы управления транскрипцией принципиально отличаются от такого варианта.
«В типичной эукариотической клетке ... транскрибируется только 7% всех последовательностей ДНК. Весьма маловероятно, чтобы остальные 93% ДНК были заблокированы десятками тысяч различных высокоспециализированных белков-репрессоров. Из соображений здравого смысла следует, что клетки высших организмов должны использовать вместо этого какие-то общие механизмы репрессии генов».
©Албертс и др., 1986
Действительно, эукариоты выработали общий механизм выключения активности генов, использующий намотку ДНК на нуклеосомы, что препятствует контакту ДНК-зависимых РНК-полимераз с достаточно длинным участком ДНК. Соответственно, эукариоты применили принципиально новый способ активирования генов, резко отличающийся от прокариотического. Основой нового механизма управления геномом стало использование волновых полей. Однако отказ от чисто химических способов управления генами не был стопроцентным — во многих случаях они используются эукариотами как дополнение к новому, физическому способу управления.
Практика акупунктуры распространила представление о существовании в организме неких «активных точек» и зон. Мы не будем эксплуатировать эту выгодную ситуацию. И без того ясно, что разные зоны организма — мышцы, костная ткань, спинномозговая жидкость и др. — настолько отличаются по своим свойствам, что химическое автоволновое поле этих зон принципиально не может быть всюду одинаковым, в том числе — по плотности энергии колебаний. Следовательно, нужно исходить из существования в организме зон с повышенной и пониженной концентрацией энергии химических волн.
Для реализации волнового механизма управления активностью геновγ
, природе пришлось снабдить геном механизмом, реагирующим на акустические колебания. Это достигнуто с помощью остроумного решения. Из белков-гистонов, обладающих повышенной основностью (щелочностью — прим. ред.)α, были созданы крупные гранулы — нуклеосомыβ, которые, благодаря основности, хорошо «прилипают» к нити дезоксирибонуклеиновой кислоты (ДНК). В спокойной среде нуклеосомы с намотанной на них нитью ДНК сами собой собираются в крупную спираль, названную соленоидом. Находясь в соленоиде, ДНК непригодна для перезаписи с нее информации на РНК, т.е. для транскрипции. Только когда акустические колебания нуклеоплазмы разрушат структуру соленоида, оторвут нить ДНК от «прилипших» нуклеосом, — вот тогда к ДНК может подойти фермент РНК-полимераза, выполняющий синтез РНК. Затем по информации РНК особые молекулярные структуры — рибосомы — соединяют нужные аминокислоты в цепочки будущих белков.[Определенный интерес в этой связи представляет собой высказывание Э. Ревича, сделанное им еще в 1986 году, цитируем:
«Первичной характеристикой рака (т.е. его первопричиной) является спонтанное образование патологических гистоновα
. Гистоны являются важнейшими элементами во всей биологии. Из гистоновα образуются гистосомы, представляющие собой сферические образования из щелочных аминокислот связанные с другими аминокислотами. Все это вместе взятое составляет основу ядерного материала каждой клетки. Связывающиеся друг с другом гистосомы образуют нуклеосомыβ, а связывающиеся друг с другом нуклеосомы образуют геныγ. Таким образом, главным причинным фактором всех раков являются патологические гистоныα, из которых формируются патологические гистосомы. Всякий процесс в организме — нормальный или патологический — запускается конкретным геномγ.Всякое заболевание подразумевает присутствие инородных гистосом. Если инородные гистосомы возникают эндогенным путем (т.е. внутри самого организма), то это называется изопаразитом. Иными словами, это примерно то же самое, как быть атакованным представителем другого биологического вида, но только этот интервент появился не извне, а образовался в собственном теле в результате мутации гистоновα
, затем гистосом, затем нуклеосомβ и затем геновγ.Итак, второй важнейший момент: рак — это изопаразит, возникший в результате повреждения или дефекта в гистонахα
, которого можно образно представлять себе в виде представителя примитивного биологического вида, размножающегося путем формирования и распространения кист. Иными словами, рак — это спонтанное возникновение правовращающих аминокислот».Довольно примечательно то, что Ревич и понятия не имел ни о какой КСГ, хотя о волновом механизме управления активностью генов, скорее всего, догадывался.]
Картина химического автоволнового поля организма закономерно отображается гетероволновой оптикой ядра каждой клетки — К-оптикой — в акустическом поле ядра. Каждой зоне повышенной плотности энергии волнового поля организма отвечает микроскопическая зона повышенной интенсивности акустических волн в ядре. Когда плотность акустической энергии превышает определенный порог, акустические колебания отрывают спираль ДНК от нуклеосом, т.е. разрыхляют хроматин, дают возможность ферментам подойти к расположенному в данном месте гену и тем создают условия для его транскрибирования. Так активные зоны химического волнового поля организма определяют в каждой клетке активность тех или иных генов.
[Хотя это не имеет прямого отношения к освещаемой теме, вероятно, все же имеет смысл вспомнить хорошо известный многим прибор Витафон, терапевтический эффект которого основан на сугубо акустическом воздействии на тело человека.]
К настоящему времени известны многочисленные механизмы управления работой генома и процессами развития, имеющие сугубо химическую природу. В последующем изложении мы не будем упоминать их потому, что они уже были объектами многих исследований, и не составляют предмета нашего рассмотрения. Но молчание по их поводу не нужно расценивать как свидетельство игнорирования этих механизмов. Напротив, мы считаем их важными компонентами информационной системы организма. Просто в проблеме формирования многоклеточного организма не они оказались главными.
Главное средство управления биохимическими процессами в клетках — локальное разрыхление хроматина — никак не удается свести к некому химическому агенту, проявляющему химические предпочтения. Оно способно затронуть любой (в химическом смысле) ген, но почему-то, в большинстве случаев, зависит от расположения клетки в организме, что совершенно нехарактерно для химических процессов. Это ярко обнаруживается при разрыхлении гена внутри сплошной массы неактивного гетерохроматина, содержащего разные гены, в том числе и точно такие же как данный.
Подобные факты говорят в пользу физической природы процесса разрыхления, безразличного к химическим особенностям генов, но учитывающего их координаты.
Ряд данных говорит не только в пользу волнового механизма разрыхления хроматина, но и в пользу определенной пространственной организации волн, например, фокусировки. Именно действие сфокусированных колебаний способно объяснить выборочное разрыхление хроматина в компактной зоне, окруженной массивом плотного гетерохроматина. В пользу пространственной организации волновых процессов, управляющих генами, говорят и закономерности пространственного расположения генов эукариот, и их пространственная фиксация.
При молекулярных размерах генов избирательное волновое управление их разрыхлением должно представлять собой непростую задачу. Трудно создать интенсивные колебания в малой зоне, не затронув соседние зоны, где расположены другие гены, не требующие активирования. Прокариотам, использующим химические, контактные способы активирования генов, легче обеспечить избирательное управление транскрипцией. Их гены могут располагаться вплотную друг к другу без опасений непроизвольного активирования соседнего гена.
Есть ли свидетельства того, что Природе было труднее обеспечить раздельное, независимое активирование генов эукариот по сравнению с прокариотами? Да, можно назвать, по крайней мере, два таких факта.
1.Во-первых, гены эукариот, в отличие от прокариот, не расположены вплотную друг к другу — между ними вставлены неинформативные спейсеры (разделители, от англ. spacer — прокладка, распорка), вдесятеро и более превышающие по длине сами гены и соответственно увеличивающие общую массу генома.
2.Во-вторых, точность управления активированием генов у эукариот значительно снижена. В отличие от прокариот, у них, хотя и с малой скоростью, синтезируются РНК почти со всех имеющихся генов.
По мнению исследователей, «...можно предположить, что слабая функция генов ... отражает ... несовершенство регуляторного аппарата, запирающего гены. Что-то вроде неплотно прикрытых «подтекающих» кранов» [Нейфах, Лозовская, 1984]. Так что признаки волнового механизма активирования генов закономерно проявились и в этом.
Характерно то, что считывание информации с ДНК происходит не только при транскрипции, но и при репликации — т.е. при удвоении числа хромосом перед делением клетки. С химической точки зрения эти процессы очень похожи друг на друга — в одном случае на матрице ДНК синтезируется нить РНК, а в другом — нить ДНК. В половине случаев (когда матрицей служит так называемая «отстающая» нить ДНК) процесс начинается совершенно одинаково — синтезируется нить РНК. В случае репликации, образовавшийся праймер (т.е. «затравка») в дальнейшем замещается цепочкой ДНК, а с 11-го нуклеотида и сам синтез цепочки нуклеотидов переключается на производство ДНК. Но, несмотря на такое сходство, репликация не зависит от разрыхления хроматина, от связи ДНК с нуклеосомами, а транскрипция — зависит.
Если для синтеза первого десятка нуклеотидов РНК не нужен отрыв ДНК от нуклеосом, то почему такой отрыв становится необходимым, когда дело доходит до синтеза РНК, несущих коды белков, причем это наблюдается у всех эукариот и, следовательно, жестко контролируется естественным отбором?
С химической точки зрения, отличие продуктов репликации (ДНК) от продуктов транскрипции (РНК) — в том, что в сахаре, содержащемся в цепочке нуклеотидов, одна гидроксильная группа заменена атомом водорода, а урацил (один из четырех нуклеотидов РНК) заменен тимином, где место одного из атомов водорода заняла метильная группа. В общей сложной структуре цепочки нуклеотидов эти различия незначительны и сами по себе не могут ответить на сакраментальный вопрос — почему, в итоге, для получения молекул РНК требуется разрыхление хроматина, а для получения ДНК оно не требуется?
Только КСГ разъясняет эту странную ситуацию. Формирование многоклеточных организмов требует учета расположения клеток в трехмерной структуре, и для управления таким процессом Природа использовала принцип оптической проекции активных зон волновых полей организма на гены. Механизм разрыхления хроматина выбран Природой как способ управления активностью генов со стороны волнового поля. Этот же способ управления обусловил удивительную консервативность аминокислотных последовательностей некоторых гистонов, без чего не было бы стабильности порогового усилия, при котором нуклеосомы начинают отрываться от ДНК.
Управление активированием генов у всех эукариот через разрыхление хроматина показывает, что, при всем многообразии химических механизмов управления внутриклеточными процессами, именно волновой механизм управления транскрипцией оказался для эукариот решающим.
Вот, почему ДНК эукариот — в отличие от прокариот — «намотана» на нуклеосомы и, соответственно, изменены ферменты, синтезирующие у эукариот новую ДНК (ДНК-полимераза) и РНК (РНК-полимеразы). Первый фермент приобрел способность работать независимо от разрыхления хроматина (такой проблемы не было у прокариот), а второй (точнее — РНК-полимеразы I и II) — только при разрыхленном хроматине. Те загадки, о которых мы говорили выше, появились как результат выработанных естественным отбором особенностей ДНК- и РНК-полимераз эукариот.
Чтобы оценить степень соответствия между следствиями КСГ и биологическими фактами, удобно одновременно сравнивать факты со следствиями теории диссипативных структур (ТДС). Поэтому отметим, что ТДС не объясняет необходимости разрыхления хроматина для синтеза РНК, притом, что синтез ДНК происходит без такого разрыхления. А для КСГ этот же факт стал одной из фундаментальных опор.
F138
--------------------------------------------------------------------------------
Исследуя механизмы шока, Ревич обнаружил потрясающий эффект: если подопытное животное ввергается в состояние сверхострого шока — наступает распространенное «продырявливание» многих частей тела.
Небольшое пояснение. Сверхострый шок у животных легко вызывается внутривенным введением коллоидного металла (см. ниже), ошпариванием горячей водой или путем нанесения тяжелой механической травмы. Для следующего вслед за этим состояния наиболее характерными и очевидными являются страшные симптомы со стороны центральной нервной системы, включая экзофтальм (выпучивание глаз) и паралич задних конечностей, за которым следуют судорожные движения, обычно с летальным исходом. Точно такой же сверхострый шок наблюдается у людей после переливания крови, несовместимой по групповому признаку, или после очень тяжелой травмы.
Чтобы детально изучить изменения, происходящие в организме вследствие сверхострого шока, Ревич «по пояс» макал подопытных крыс в кипяток, после чего извлекал из них разные части тела и проводил гистологический анализ. Так вот самым характерным нарушением при сверхостром шоке оказалась клеточная вакуолизация, т.е. появление «дырки» внутри изначально целостной клетки. Такие вакуоли присутствуют в клетках паренхимы печени, в меньшей степени — в альвеолярных клетках легкого, и еще в меньшем количестве — в клетках почек. Но особый интерес вызывает обнаружение вакуолей в цитоплазме и даже в ядрах клеток головного мозга — именно этот факт объясняет манифестацию при сверхостром шоке тяжелых неврологических симптомов.
При менее тяжелых формах шока вакуолизация клеток также имеет место быть, но в существенно меньшем объеме — главные патологические изменения затрагивают мышцы. Кроме всего прочего, эти изменения проявляются сосудистой и интерстициальной патологией, такой как отеки и капиллярные кровотечения. Особыми характеристиками состояния обычного (не сверхострого и не острого) шока являются два других специфических патологических проявления: а) милиарные поражения слизистой оболочки желудка, ведущие к кровотечению и изъязвлению, и б) явное накопление жидкости в первой части тонкого кишечника.
Возникает закономерный вопрос:
Каким образом в изначально целостных клетках без какого либо нарушения их оболочки могут внезапно массовым порядком возникать полости?
Разве не линзоподобной фокусировкой химических волн на клеточных оболочках с концентрацией преломленного энергетического пучка внутри клетки, происходящей в экстремальных обстоятельствах, вызываются эти чрезвычайно странные явления? Кстати, если положить сырое яйцо в микроволновую печь, то что будет?
Изучая изменения, происходящие в клетках вследствие облучения, Ревич обнаружил, что после облучения — особенно при использовании высоких доз — также наблюдается интенсивная клеточная вакуолизация. С помощью радиоактивного натрия было также установлено, что после облучения наблюдается большая склонность этого, по большей части внеклеточного элемента проникать в клетки.
Вакуоли
Но дальше все оказалось еще интереснее — введение в организм конъюгированных жирных кислот приводило к появлению вакуолей не только в цитоплазме, но даже в ядрах клеток! Иными словами, воздействие на клетки конъюгированных жирных кислот практически повторяет таковое радиации.Эффект от препаратов Ревича на основе селена (Se) на клеточном уровне виден даже при использовании дозировки в микрограммах — в цитоплазме клеток происходит вакуолизация! Интересно также отметить в этой связи, что, несмотря на вакуолизацию клеток, наблюдаются также и околоклеточные отеки (рис. 1).
Вакуоли в эритроцитах Обработка in vitro эритроцитов конъюгированными жирными кислотами — особенно триенами — вызывает выраженную, хорошо заметную вакуолизацию (рис. 2).
А вот в присутствии антагонистов жирных кислот, к которым относятся некоторые спирты, во многих случаях предотвращаются и вакуолизация, и даже изменения в ядрах, наблюдающиеся у облученных животных контрольной группы. Кстати сказать, кому неизвестно хоть и слабое, но все же реально существующее противолучевое действие этилового спирта?!
Стоит также упомянуть и о двух других, чрезвычайно интересных опытах Ревича.
1.Были приготовлены 1/10-молярные растворы с рН=8.5 двухосновных фосфатов лития (Li), натрия (Na), калия (K), аммония (NH4) и рубидия (Rb). Каждый из растворов вводили внутривенно мышам по ¼ мл в минуту — вплоть до момента смерти животного. После этого их органы — особенно головной мозг — немедленно фиксировались в жидкости Буэна и изучались гистологически. В случаях аммония и рубидия наблюдали присутствие вакуолей в клетках и, особенно, в ядрах! При введении калия вакуоли появлялись в цитоплазме, а после применения натрия отмечен только лишь околоклеточный отек. При введении лития никаких изменений не отмечено.
2.Лейкоциты с колларголомНормальные лейкоциты
В экссудат, полученный путем плевральной пункции и богатый гранулоцитами, добавляли небольшое количество коллоидного серебро-белкового препарата (0.1% Колларгола) и содержали при температуре +38°C. Было установлено, что, вслед за фагоцитозом крупинок серебра, это вызывало появление вакуолей в лейкоцитах. Причем вакуоли быстро увеличивались в размерах с последующим взрывом лейкоцитов! Ядра же оставались в виде ядерных «теней». На рисунке показаны нормальные гранулоциты и, рядом, вакуолизированные гранулоциты на грани взрыва с поглощенными крупинками колларгола. А теперь самое интересное: добавление минимальных количеств морфина или иных дериватов опия полностью предотвращает эти изменения в лейкоцитах — вакуоли не обнаруживаются и лейкоциты не взрываются. Причем оказалось, что способностью предотвращать взрыв лейкоцитов обладают только морфин и прочие опиумные алкалоиды. Ни адреналин, ни хинин, ни прочие исследованные вещества никакого эффекта не оказывают.
www.ling1.net/
@темы: Равновесие, Вопрос, Соль, Физическое тело